製品・サービス情報
|
|
ArrayStar社 遺伝子発現制御機構解析
non-coding RNA マイクロアレイ受託解析サービス |
|
 |
 GlycoRNA 研究 GlycoRNA 研究
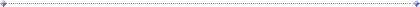
|
・GlycoRNA とは?
・なぜ GlycoRNA を研究するのか
・GlycoRNAの 研究方法
◆GlycoRNA とは?
|
glycoRNA は、糖鎖付加を受けた低分子ノンコーディング RNA で、核内低分子 RNA (snRNA)、リボソーム RNA (rRNA)、核小体低分子
RNA (snoRNA)、トランスファー RNA (tRNA)、Y-RNA[1]、micro RNA[2] などが含まれます。グリコシル化 mRNA
は知られていません。特に glycoRNA 上の N-グリカンは、シアル化とフコシル化が進んでいます。glycoRNA は細胞表面に存在し、免疫応答の調節に重要な
Siglec 受容体に結合することができます (図1)。
脂質とタンパク質は糖で修飾された唯一の生体高分子であると考えられており、RNA >がグリコシル化されるとは考えられていませんでした。2021
年、ノーベル賞受賞者のベルトッツィ研究室で、糖鎖前駆体の代謝標識により哺乳類の低分子ノンコーディング RNA がシアリル化糖鎖と結合することが発見されました。従来の常識は覆され、RNA
および糖鎖生物学の分野で大きな進歩を遂げました [1]。
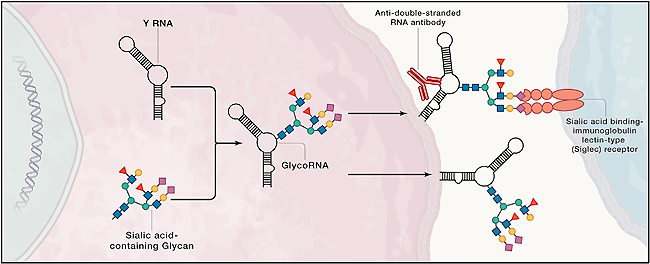
図1.glycoRNA は、シリアル化糖鎖による RNA の糖鎖修飾によって生成され、細胞膜上に表示され、シアル酸結合免疫グロブリン型レクチン(Siglec)受容体によって認識される[1,
5]。
RNA のグリコシル化の基礎となるメカニズムはまだ明らかになっていません。glycoRNA の生合成は、既存の標準的な N グリカン生合成機構に依存しています[1]。ldlD変異体 CHO 細胞および CRISPR-GALE ノックアウト K562 細胞は、N グリカン生合成経路に欠損があり、glycoRNA の産生が阻害されています。外部のグリカンを導入により、この阻害を逆転させることができます。さらに、タンパク質の N-グリコシル化を媒介する酵素であるオリゴ糖転移酵素 (OST) の阻害は、glycoRNA 産生の減少を引き起こします。糖タンパク質中の N および O 結合型グリカンの場合、糖は主に小胞体内で付加され、その後ゴルジ体でさらに修飾された後、輸送または分泌のための小胞に最終的に包装されます。しかしながら逆説的なことに、RNA は通常これらの細胞コンパートメントには存在しません。まとめると、RNA のグリコシル化は、少なくとも部分的にはタンパク質のグリコシル化と同じ機構とメカニズムを使用します。一方、独立した経路を考慮しないわけではありません。
・グリコシル化結合
|
糖鎖-RNA の化学結合の正確な性質は、現在活発に研究されています。glycoRNA は、有機相分離、プロテイナーゼ K 処理、シリカベースの
RNA 精製、高濃度のホルムアミドでの加熱などの厳しい変性条件に耐えられるほど頑強であるため、非共有結合である可能性は低いです。glycoRNA
は、糖タンパク質中の N-グリカンのアスパラギンと近位 GlcNAc との間の結合を切断する PNGase F に感受性があります。しかしながら、天然の
RNA 核酸塩基は、このようなアミド結合を含むリンカーがありません。糖タンパク質と同様の RNA-糖鎖結合を確立するためには、RNA における追加のグアノシン修飾が必要です。もう一つの可能性は、RNA
とあらかじめ会合した N-グリカンが、ペプチド性または非ペプチド性の小さなリンカーによって結合されていることです。RNA-糖鎖結合の化学的および構造的性質を定義することは、今後の
glycoRNA 研究においてきわめて重要でしょう。 RNA 最適化過ヨウ素酸酸化およびアルデヒドライゲーション(rPAL)および >sequential
window acquisition of all theoretical mass spectra (SWATH-MS) により、修飾された
RNA3-(3-アミノ-3-カルボキシプロピル)ウリジンにおけるN-N グリカンの主要な結合部位であることが判明しました[7]。
図2.修飾 RNA 塩基 acp3U を介した N グリカンと RNA の化学結合[7]。
|
・GlycoRNA の機能と疾患
|
glycoRNA は、細胞分画や免疫組織化学イメージングで示されるように、主に細胞表面に存在し、シアル酸切断酵素処理によって細胞表面から除去することができます[1]。細胞表面への局在は、細胞外相互作用を媒介する役割を示唆しています。特に、glycoRNA
はシアル酸結合免疫グロブリン様レクチン (Siglec) 受容体ファミリーと結合親和性を有しており(図1)、標的細胞との細胞接着、細胞シグナル伝達、免疫応答調節における直接的なリガンドとしての関与が強調されています[1]。さらに、Siglec
受容体ファミリーのリガンドパートナーは、これまでの研究が糖脂質と糖タンパク質のみに限定されていたこともあり、ほとんど知られていません。新たに発見された
glycoRNA は、これらの Siglec 受容体や他のオーファン糖鎖結合受容体の、長い間探し求められていたリガンドである可能性があります[5-6]。
タンパク質および脂質のグリコシル化の異常は、長い間、さまざまなヒトの疾患の特徴として確立されてきました。同様に、glycoRNA も、疾患において糖鎖ネットワークを破壊する可能性を秘めています。例えば、低分子ノンコーディング
glycoRNA の中でも、Y-RNA は特に興味深い存在で、その結合タンパク質やリボ核タンパク質 (ribonucleoproteins:
RNP) は、全身性エリテマトーデス (systemic lupus erythematosus: SLE) などの自己免疫疾患に関連する抗原として知られています[1]。その他の
glycoRNA は、癌、心血管疾患、神経疾患、免疫疾患、呼吸器疾患に関与していることがわかっています。
治療研究の新たな道筋として、glycoRNA糖鎖の修飾が免疫療法薬に対する応答性に影響を与える可能性があります。RNA 配列部分を標的とするアンチセンスは、合理的な薬物設計に適しており、高い選択性が期待できます。特に、高分子治療モダリティは、細胞外からアクセスして細胞表面に露出した標的
glycoRNA と反応することができ、細胞内に侵入する困難さはありません。バイオマーカー用途では、糖鎖および RNA 部分は、免疫化学的および塩基配列に基づく検出法の両方に対応する生化学的特性を有しているため、高感度で特異的な疾患の診断と予後予測が可能になります
[3-4]。
新しい生体分子クラスとしての glycoRNA は、トランスクリプトミクス、エピトランスクリプトミクス、RNA 生物学、糖鎖生物学、細胞生物学、生化学、シグナル伝達、免疫学、基礎生物学および生物医学/臨床科学において変革の可能性を秘めています。Arraystar
GlycoRNA Array は、広範な科学研究分野や多くの臨床疾患における glycoRNA をプロファイリングおよび同定が可能です。
|
【Reference】
| 1. |
Flynn RA. et al (2021) "Small RNAs are modified
with N-glycans and displayed on the surface of living cells." Cell
184(12):3109-3124.e22 [PMID:34004145] |
| 2. |
Li, J. et al (2024) "O-Glycosylated RNA
Identification and Site-specific Prediction by Solid-phase Chemoenzymatic
TnORNA method and PONglyRNA tool" bioRxiv [doi: https://doi.org/10.1101/2024.06.18.599663] |
| 3. |
Angata K. et al (2020) "Glycogene Expression
Profiling of Hepatic Cells by RNA-Seq Analysis for Glyco-Biomarker
Identification." Front Oncol 10:1224 [PMID:32850363] |
| 4. |
Nishimura S (2011) "Toward automated glycan
analysis." Adv Carbohydr Chem Biochem 65:219-71 [PMID:21763513] |
| 5. |
Disney MD (2021) "A glimpse at the glycoRNA
world." Cell 184(12):3080-3081 [PMID:34115968] |
| 6. |
Clyde D (2021) "Sugar-coated RNAs." Nat
Rev Genet 22(8):480 [PMID:34168329] |
| 7. |
Xie Y. et al (2024) "The modified RNA base
acp(3)U is an attachment site for N-glycans in glycoRNA." Cell
187(19):5228-5237.e12 [PMID:39173631] |
|
◆なぜ GlycoRNA を研究するのか
|
glycoRNA は、癌、心血管疾患、神経疾患、免疫疾患、呼吸器疾患における新たな研究分野として浮上し、新しいバイオマーカーおよび治療応用への新たな道を切り開いています。
・乳癌
|
表面 glycoRNA は腫瘍の悪性度や転移と逆相関しています。つまり乳癌以外の細胞 (MCF-10A) の glycoRNA レベルが最も高く、次いで乳癌細胞 (MCF-7)、一方、乳房転移癌細胞 (MDA-MB-231) のレベルは最も低くなります (図1)。この逆相関は、これらの glycoRNA が乳がんの進行と転移のバイオマーカーとして利用できることを示唆しています。
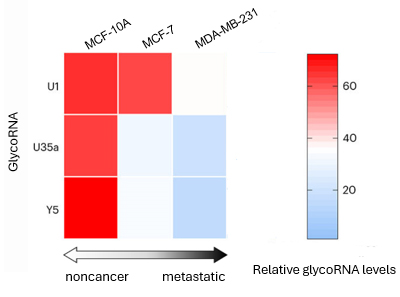
図1.シアル酸アプタマーと RNA in situ ハイブリダイゼーションを介した近接ライゲーションアッセイ(aptamer and RNA
in situ hybridization-mediated proximity ligation assay:ARPLA)により測定された、様々な悪性度および転移段階の乳癌細胞における
glycoRNA レベルのヒートマップ。 |
・膵臓癌
|
特定のグリコシル化 microRNA は、膵臓癌細胞の成長と増殖の調節因子として同定されています[2]。例えば、グリコシル化 miR-103a-3p、miR-122-5p、miR-4492
は、PI3K-Akt 経路を通じて膵臓癌細胞の成長と増殖を調節することができます[2-3](図2)。さらに、グリコール-miRNA(例えば has-miR-21-5p)
の発現に影響を与えることにより、β-1,4-ガラクトシルトランスフェラーゼ 1(B4GALT1) は、MIA PaCa-2 膵臓癌細胞の細胞周期を抑制し、アポトーシスを促進することができます。したがって、B4GALT1
miRNA のグリコシル化酵素である可能性があります[3]。
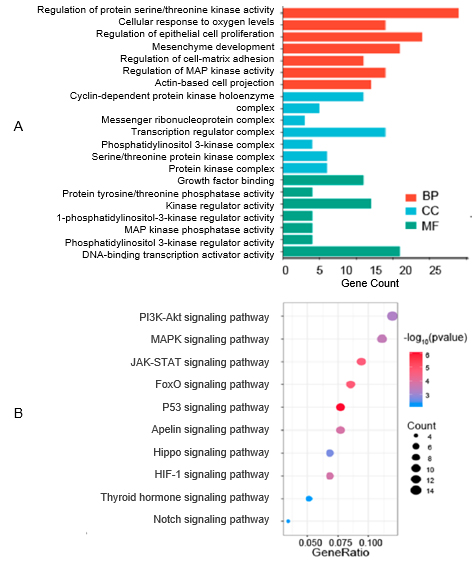
図2.Gene Ontology(A) および KEGG パスウェイ(B) 解析において、膵臓癌細胞で差次的に発現する glyco-miRNA
は、発がん性シグナル伝達経路の制御に富んでいることが示されている[3]。 |
・心血管疾患
|
Y-RNA は、高度に保存された低分子のノンコーディング RNA (~110 nt) の一種で、グリコシル化の主要な標的です [6-7]。Y-RNA
は免疫調節において重要な役割を果たしています。興味深いことに、Y-RNA はヒトの循環 RNA の 60% 以上を占めており、そのレベルは虚血性脳卒中の主要な危険因子であるアテローム性動脈硬化症の進行と正の相関があります
[4-5]。具体的には、Y-RNA フラグメントはマクロファージを活性化し、アテローム性動脈硬化症において炎症を促進します [5]。これらの観察結果は、グリコシル化
Y-RNA が心血管疾患の危険因子およびバイオマーカーとなる可能性があることを示唆しています。 |
・脳卒中
|
glycoRNA は、虚血性脳の炎症反応を調節すると考えられています[10](図
)。神経細胞の糖タンパク質と同様に、glycoRNA はミクログリアの Siglec-11 受容体のリガンドとして作用し、IL-1b のようなサイトカインや一酸化窒素を生成する NOS2 などの発現を減少させることで、ミクログリアを抗炎症表現型に偏向させる可能性があります。
glycoRNA 経路を標的とすることは、脳卒中の治療と回復のための有望なアプローチとなる可能性があります。
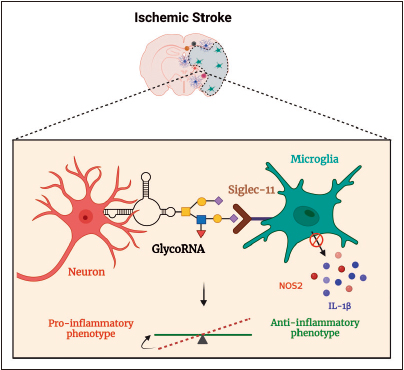
図3.脳卒中後の糖鎖 RNA の推定的役割。神経細胞表面の glycoRNA
は、ミクログリアの Siglec-11 受容体のリガンドとして機能し、IL-1b サイトカインと一酸化窒素の産生を減少させ、虚血性脳卒中後に抗炎症カスケードを開始する [10]。 |
・免疫調節
|
glycoRNA は、免疫細胞受容体、特にシアル酸結合免疫グロブリン様レクチン (Siglec) 受容体ファミリーのメンバーと相互作用することがわかっています[6]。好中球表面の glycoRNA は、好中球の内皮細胞への接着制御、経内皮移動の促進、好中球動員の制御に必須です[8](図4)。さらに、レクチンSelpは、Seleとは対照的に、好中球 glycoRNA の少なくともサブセットを同定することができます。したがって、Selpの glycoRNA リガンドは、非常に類似したレクチンタンパク質の間でも、独自のリガンド特異性を付与する可能性があります[8]。免疫細胞上の
glycoRNA は、さまざまな免疫関連疾患や自己免疫疾患に広く関与している可能性があります。
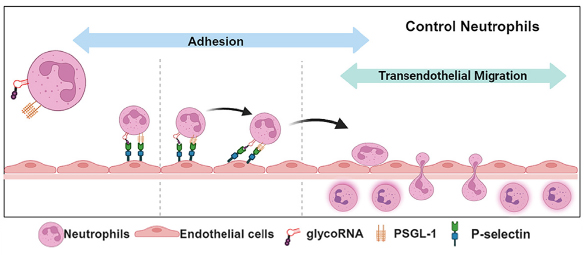
図4.glycoRNA は、p-セレクチンのリガンドとして機能し、好中球の動員を制御する。 |
・肺胞上皮機能
|
glycoRNA は、肺胞上皮グリコカリックスの新規成分であり、上皮バリア機能を制御したり、インフルエンザ A ウイルス感染に影響を与えたりします[9]。マクロファージ、単球、好中球などの免疫細胞上のシアル酸結合免疫グロブリン様レクチン (siglecs)11 および 14 のリガンドとしての glycoRNA の同定された機能は、炎症性肺疾患における免疫細胞の相互作用に影響を与える可能性があります[6]。さらに、肺胞上皮細胞の表面に存在する glycoRNA は、サーファクタントプロテイン A や D などの肺コレクチンと結合し、呼吸器免疫応答にさらに寄与する可能性があります[9]。したがって、気管支肺胞洗浄液中に排出された glycoRNA の存在を分析することは、上皮の損傷と破壊を評価するための貴重なバイオマーカーとなる可能性があります。
|
【Reference】
| 1. |
Ma Y. et al (2024) "Spatial imaging of
glycoRNA in single cells with ARPLA" Nat Biotechnol 42(4):608-616
[PMID:37217750] |
| 2. |
Li, J. et al (2024) "O-Glycosylated RNA
Identification and Site-specific Prediction by Solid-phase Chemoenzymatic
TnORNA method and PONglyRNA tool" bioRxiv [doi: https://doi.org/10.1101/2024.06.18.599663] |
| 3. |
Li J. et al (2023) "Novel Approach to
Enriching Glycosylated RNAs: Specific Capture of GlycoRNAs via Solid-Phase
Chemistry." Anal Chem 95(32):11969-11977 [PMID:37524653] |
| 4. |
Driedonks TAP. and Nolte-'t Hoen ENM. (2018)
"Circulating Y-RNAs in Extracellular Vesicles and Ribonucleoprotein
Complexes; Implications for the Immune System." Front Immunol 9:3164
[PMID:30697216] |
| 5. |
Hizir Z. et al (2017) "RNY (YRNA)-derived
small RNAs regulate cell death and inflammation in monocytes/macrophages."
Cell Death Dis 8(1):e2530 [PMID:28055017] |
| 6. |
Flynn RA. et al (2021) "Small RNAs are
modified with N-glycans and displayed on the surface of living cells."
Cell 184(12):3109-3124.e22 [PMID:34004145] |
| 7. |
Flynn, RA. et al (2019) "Mammalian Y RNAs are
modified at discrete guanosine residues with N-glycans." bioRxiv [https://doi.org/10.1101/787614
] |
| 8. |
Zhang N. et al (2024) "Cell surface RNAs
control neutrophil recruitment." Cell 187(4):846-860.e17 [PMID:38262409] |
| 9. |
Abledu, JK. et al (2024) "Cell surface RNA
expression modulates alveolar epithelial function." bioRxiv [https://doi.org/10.1101/2024.05.19.594844] |
| 10. |
Chokkalla AK. et al (2023) "Immunomodulatory
role of glycoRNAs in the brain." J Cereb Blood Flow Metab 43(4):499-504
[PMID:36644904] |
|
◆GlycoRNA の研究方法
|
・GlycoRNA Array プロファイリング
|
Arraystar社の GlycoRNA
Array は、glycoRNA の生化学的捕捉法とマイクロアレイによる RNA 検出法を組み合わせて、glycoRNA 発現の定量およびプロファイリングを行います。これら 2 つの高度な技術の統合により、両方法の長所が活用され、高い特異性、感度、精度を実現します。
本アレイは、Y-RNA/Y-RNA フラグメント、tRNA、tsRNA(tiRNA および tRF)、pre-miRNA、miRNA、snRNA/snRNA フラグメント、snoRNA/snoRNA フラグメント、rRNA/rRNA フラグメント、
scRNA など、幅広いグリコシル化 small RNA クラスをカバーしています。
この最先端のアプローチにより、研究者は包括的な glycoRNA 発現の詳細を得ることができ、遺伝子制御、細胞機能、ヒト疾患におけるこの新しいクラスの
RNA 分子を発見し理解することができます。 |
・GlycoRNA 検出法
|
非常に特異的な生化学的方法により、Total RNA サンプルから glycoRNA を検出、捕捉、濃縮することで、真の glycoRNA シグナルを確保し、検出感度を高めることが可能です。
<代謝標識とクリックケミストリー>
|
glycoRNA は、クリックケミストリー[1](図1)を用いて代謝標識されます。この方法では、アジド基を持つ N-アジドアセチルマンノサミン
(Ac4ManNAz) などのレポーター糖が細胞や組織のグリカンに取り込まれます。次に、単離された RNA のグリカンを、アジドとジベンゾシクロオクチン-ビオチン
(DBCO-ビオチン) などの標識との間に結合させ、アジド-アルキン環化付加クリックケミストリーによって結合します(図5)。
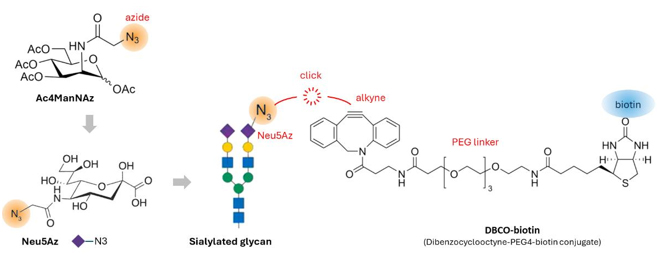
図5.Ac4ManNAz(N-アジドアセチルマンノサミン-テトラアシル化)は、Neu5 シアル酸 (N-アセチルノイラミン酸
)の前駆体である。細胞培養中に細胞透過性を高めるために、ペルアセチル化される。細胞内では、Ac4ManNAz は代謝的に Neu5Az に変換され、シアル化糖鎖の一部となる。
In vitro では、Neu5Az 中のアジドと
DBCO-ビオチンクリック試薬のアルキンが銅フリーのクリックケミストリーによって結合し、グリカンをビオチンで標識することができる。 |
<過ヨウ素酸酸化およびアルデヒドライゲーション法(Periodate Oxidation and Aldehyde Ligation: pAL)>
|
過ヨウ素酸酸化およびアルデヒドライゲーション法 (pAL) は、生細胞での代謝標識を必要とせずにネイティブな glycoRNA を標識するように設計されています[2]。この方法では、過ヨウ素酸塩を用いて、glycoRNA
のシアル酸ジオールを最適な条件下で選択的にアルデヒド基に酸化し、その後、アルデヒド反応性ビオチン試薬のアミン基とアルデヒドライゲーションを行います(図6)。rPAL
は堅牢で柔軟性があり、従来の sialoglycoRNA 検出法よりも感度が少なくとも一桁高い感度を備えています。

図6.RNA 過ヨウ素酸酸化およびアルデヒドライゲーション(rPAL)では、シアル酸ジオールを過ヨウ素酸によってアルデヒドに酸化し、ビオチンなどのアルデヒド反応性標識のアミン基にライゲーションすることができる。 |
<生体内細胞過ヨウ素酸酸化およびアルデヒド標識(pAL)>
|
glycoRNA の生体内細胞過ヨウ素酸酸化およびアルデヒド標識 (pAL) は、生細胞表面および細胞膜関連 RNA の glycoRNA を蛍光色素またはビオチンで標識するために使用されます [3]。具体的には、過ヨウ素酸ナトリウムを使用して軽度の酸化処理を施した細胞表面 glycoRNA に Cy5-ヒドラジドまたはビオチン-ヒドラジドを結合させます。
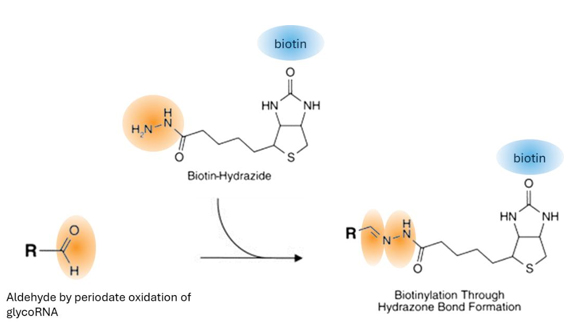
図7.glycoRNA 中のシアル酸ジオールは過ヨウ素酸とともに酸化され、アルデヒド基を形成する。 次に、ビオチンまたは
Cy5 ヒドラジドは、アルデヒド基を介して glycoRNA に結合する。 |
<GlycoRNA の細胞表面イメージング>
|
細胞表面上における glycoRNA の発現と局在は、例えば、シアル酸アプタマーと RNA in situ ハイブリダイゼーション媒介近接ライゲーションアッセイ(ARPLA)[4]
によって視覚的に画像化することができます。図8 に示すように、ARPLA は以下の機能要素から構成されます:(1)N-アセチルノイラミン酸(Neu5Ac)
と選択的に結合する「アプタマー」、ハイブリダイゼーション中の立体障害を回避する「スペーサー」、およびその後の近接ライゲーションのための DNA
リンカー (「リンカー G」) を有する糖鎖プローブ; (2)RNA in situハイブリダイゼーション「RISH」用の DNA 鎖、もう一つの「スペーサー」、およびリンカー
G と協働する DNA リンカー (「リンカーR」) を有する RNA 結合プローブ; (3)RCA 鋳型として環状 DNA を生成するために
in situ ライゲーションを可能にする、リンカー G および R とハイブリダイズするコネクター 1 および 2;(4)glycoRNA
検出シグナルを報告するための RCA 産物に相補的な蛍光色素結合一本鎖 DNA (ssDNA)プローブからなるレポーター。二重のグリカン-RNA
結合と in situ ライゲーションにより、高い選択性と低い偽陽性が保証されます。ローリングサークル増幅は、非常に感度の高感度のシグナルが生成されます。
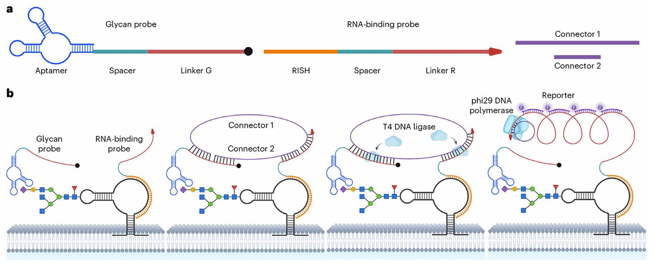
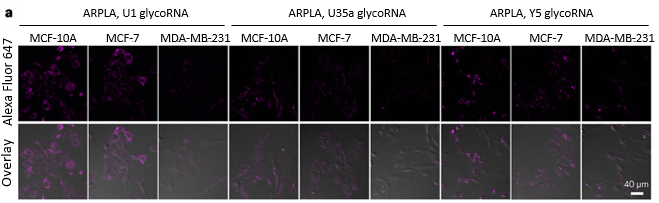
図8.ARPLA を用いた glycoRNA イメージング
(a) 糖鎖プローブ、RNA 結合プローブ、およびコネクターの概略図。
(b) これらの glycoRNA への同時結合により、
T4 DNA リガーゼのライゲーションが起こり、phi29 DNA ポリメラーゼを用いたローリングサークル増幅
(RCA) の鋳型が形成される。増幅された RCA 産物は、蛍光レポータープローブのハイブリダイゼーションによって検出される。
(c)MCF-10A、MCF-7 および MDA-MB-231 細胞における
U1、U35a、Y5 glycoRNA の ARPLA イメージング。 |
<レクチン親和性結合>
|
コムギ胚芽凝集素 (WGA) や Maackia amurensis Lectin II(MALII) などのレクチンは、glycoRNA 上に存在するシアリル化糖鎖構造を認識し、直接結合します[1]。ビオチン-レクチン親和性結合に基づくglycoRNA 精製は、グリコシル化 RNA を単離し濃縮する効果的な方法です。
|
|
・バイオインフォマティクスツール
|
バイオインフォマティクスツールは glycoRNA の研究の大きな助けとなります。例えば、GlyinsRNA は、RNA 分子上の推定グリコシル化部位を予測することができます[5]。GlyinsRNA はグリコシル化および非グリコシル化の small RNA 配列 [6] を使用して、機械学習モデルをトレーニングします。GlyinsRNA は、オンラインのウェブサーバーとして実装されており、予測されたグリコシル化部位と過剰発現した RNA 結合タンパク質 (RBP) 関連モチーフの両方にアノテーションを付加します。 |
・機能研究
|
<GlycoRNA 結合タンパク質>
|
glycoRNA と結合するパートナータンパク質は、glycoRNA がどのような機能を持つかを示す優れた指標となります。相互作用するタンパク質は、グリカンに結合するもの
(Siglec など) と RNA 部分に結合するもの (RBP) があります。例えば、glycoRNA の免疫調節能を調べるために、glycoRNA
とシアル酸結合性免疫グロブリン様レクチン(Siglec) 受容体ファミリーのメンバーとの相互作用を免疫学的アッセイで評価することができます[1,
7]。これらの相互作用を調べることで、科学者は、glycoRNA が免疫応答にどのような影響を及ぼし、様々な疾患プロセス、特に神経炎症や自己免疫疾患の状況にどのように関与しているのかを知ることができます
[1、7]。逆に、glycoRNA は、未知の同族リガンドを持つ多くのオーファン受容体のリガンドとしてスクリーニングできます。 |
<遺伝学的、薬理学的、酵素学的アプローチ>
|
glycoRNA の生合成と機能の研究には、遺伝的阻害アプローチと薬理学的阻害アプローチの両方が用いられてきました。例えば、UDP-グルコース(Glc)/GlcNAc を UDP ガラクトースに相互変換する能力を欠く遺伝的 ldlD 変異細胞株、UDP-ガラクトース-4-エピメラーゼの CRISPR-Cas9 ノックアウト、および糖鎖生合成薬理学的阻害剤 (NGI-1、Kifunensine、Swainsonine など) が、glycoRNA 生合成
における糖鎖生合成機構の観察に用いられてきました[1]。シアリダーゼ、
PNGaseF、Endo-F2、-F3、
-Hf、 O-デグリコシダーゼなど、糖鎖構造に対する特異性の異なるエンドグリコシダーゼのパネルを使用して、糖鎖構造と
glycoRNA 機能を解析することができます[1]。このような研究は、
glycoRNA 形成の基礎となるメカニズムと、様々な細胞プロセスや病態における glycoRNA の潜在的な役割について貴重な知見をもたらしました。
|
|
【Reference】
| 1. |
Flynn, R.A., et al. (2021) "Small RNAs are
modified with N-glycans and displayed on the surface of living cells" Cell
184(12):3109-3124 e22 [PMID:34004145] |
| 2. |
Hemberger, H., et al. (2023) "Rapid and
sensitive detection of native glycoRNAs" bioRxiv 2023.02.26.530106 |
| 3. |
Abledu, J.K., et al. (2024) "Cell surface RNA
expression modulates alveolar epithelial function" bioRxiv
2024.05.19.594844 |
| 4. |
Ma, Y., et al. (2024) "Spatial imaging of
glycoRNA in single cells with ARPLA" Nat Biotechnol 42(4):608-616
[PMID:37217750] |
| 5. |
Cui, C., et al. (2021) "GlyinsRNA: a webserver
for predicting glycosylation sites on small RNAs" RNA Biol
18(sup2):600-603 [PMID:34559595] |
| 6. |
Flynn, R.A., et al. (2019) "Mammalian Y RNAs
are modified at discrete guanosine residues with N-glycans" bioRxiv
787614 |
| 7. |
Zhang, N., et al. (2024) "Cell surface RNAs
control neutrophil recruitment" Cell 187(4):846-860 e17 [PMID:38262409] |
|
|
*本サイトの情報は、Arraystar社のHomepageの情報を一部引用しております。
|
|
